Featured Research

The Arboretum grounds are used by many researchers, including undergraduate and graduate students, faculty, and researchers from external organizations. If you would like further information on research at The Arboretum please contact our Research Coordinator, Dr. Aron Fazekas at afazekas@uoguelph.ca.
Genotype vs local environmental factors in regulating leaf abscission in temperate deciduous trees
By McKayla Jarvie, Catherine Rutherford, Katrina Mamone and Bryan Oblin; Undergraduate students at the University of Guelph
Climate change, caused by various human activities, continues to alter biological systems across the world, potentially interfering with important ecosystem services such as water and carbon cycling that humans rely on. Autumn leaf abscission (leaf drop) in trees is crucial for these environmental cycles to take place as both water and carbon are absorbed and released as seasons change. With these services being so vital, it is more important than ever to understand how deciduous tree species will respond to the significant changes in climate expected in the next few decades. During the Lab & Fieldwork in Ecology course, we tested whether genetics or local environmental conditions were the primary control of abscission. It is possible, that individuals from different latitudes show different abscission timing as a result of natural selection on genetic components that control the timing of abscission. Alternately, it is also possible that individuals of a species display the same abscission timing as a result of similar temperatures and levels of precipitation experienced in a particular location (the Arboretum) regardless of their latitudinal origin. For our research project, we selected various individual trees from three native species, (red maple, sugar maple and the threatened blue ash) in the arboretum’s Gene Bank and other collections and grouped them based on the latitude from which they were originally collected. Beginning in October, the abscission of each individual tree was assessed every other day based on the percentage of leaves it had lost until the tree was bare.

A red maple individual whose latitudinal origin was near Sault Ste. Marie. The picture on the left was taken on October 12, 2018, and shows full foliage while the picture on the right was taken on October 26, 2018 with the majority of leaves having fallen.
We had expected one of two things to occur: either abscission timing would be the same for all trees, indicating that the local environment was the primary influence, or timing would be different, varying according to latitude, indicating that genetics had the major influence on abscission. However, our results showed no differences in abscission timing between latitude origins, but rather a variation in timing existed within latitude groups instead. Seeing as these conflicting results were not expected, it led us to new hypotheses for how abscission is controlled. This study was conducted with the assumption that abscission timing has one primary influence, while our results imply that this process is more likely to have two or more contributing influences.
While this research didn’t provide exact answers, it provided inspiration for future projects, including observing abscission over several years to allow for possible patterns to be revealed. This knowledge could be applied to conservation and management techniques as it may make processes such as abscission more predictable in a changing climate.
Adaptation to temperature and precipitation in petiole length of Kentucky coffeetree (Gymnocladus dioicus)
By Kayleigh Hutt-Taylor, Cameron Azulay, Katelyn Hill and Rachel Young
The evolutionary significance of plant morphological structures is fundamental in understanding species capacity to adapt to changes in the environment. Plant morphologies can arise through an evolutionary response to the local environment (local adaptation) but specific forms are often expressed in response to temporary changes in the environment. The capacity for a species to express different forms depending on environmental factors is known as phenotypic plasticity. For example, the Hawaiian tree Metrosideros polymorpha can display differing leaf hypodermis thickness depending on moisture availability. Considering how these plant structures relate to species success can aid in managing threatened populations and contribute to effective conservation strategies. The Kentucky Coffee tree (Gymnocladus dioicus), an early successional species, represents a species currently facing threats to survival that are related to its environment. The species maintains small scattered populations across Southern Ontario and the Eastern United States but faces strong dispersal barriers and isolation from other populations. These populations have experienced different abiotic conditions of temperature and precipitation. These differences in environmental conditions, combined with population isolation have created the ideal conditions for local adaptation to its geographic origin.

Our study examined whether variation in petiole length in G. dioicus individuals in the Arboretum gene bank was a result of local adaptation to levels of temperature and precipitation at the geographic origin of the individuals. Few studies have considered whether leaf petiole length adapts to the local environment to maximize sun exposure and minimize water loss. Our study aimed to investigate whether there was any adaptive significance to this plant characteristic. The Arboretum’s gene bank and ‘World of Trees’ collection offered a unique opportunity to investigate the morphological differences in petiole length in a ‘common garden’ setting. Seed from distinct geographic populations across North America have been grown at the University of Guelph’s Arboretum. This means environmental conditions such as temperature and precipitation are the same and any differences observed can be attributed to local adaptation (genetic differences) rather than phenotypic plasticity. Individuals originating from fourteen different locations across Canada were sampled to determine whether differences in petiole length could be explained by the temperature and precipitation levels of individual geographic origin.
Overall, our findings indicated no significant relationship between petiole length and climatic conditions of geographic origin and therefore could not support our hypothesis that G. dioicus individuals had experienced local adaptation in petiole length. Although local adaptation to native temperature and precipitation levels could not be confirmed in this study, further research examining G. dioicus response to changes in the environment could help determine its capacity for adaptation. G. dioicus is currently listed as a threatened species in Canada and continues to experience increased isolation and barriers to dispersal. Understanding the mechanisms related to the success of this species will be crucial in creating management strategies to promote its survival in a changing climate. Although it is unclear whether G. dioicus individuals experienced local adaptation or alternatively, experienced a combination of factors related to natural selection and phenotypic plasticity linked to their geographic origin, understanding the mechanisms involved in this species response to climatic conditions remains crucial. Further study could help close the gap of knowledge regarding its capacity to adapt to new and changing environments in the future.
Effects of elevation and Eurosta solidiginis parasitism on Solidago altissima performance in the University of Guelph Arboretum
By Savanah Muller, Cassidy Mccarthy, Aiden Maloney and Sarah Hasenack
Climate change has been shown to cause increased fluctuations of average rainfall, resulting in increasing occurrence of droughts in the Northern hemisphere. The effects of drought on plants undergoing simultaneous biotic stress, such as parasitism, could influence their internal allocation of resources and their ability to withstand such disturbances. Our study, conducted at the University of Guelph Arboretum, focused on the performance of late goldenrod (Solidago altissima) undergoing simultaneous parasitism by goldenrod gall fly (Eurosta solidiginis).

Performance of goldenrod was determined at low, mid and high elevation plots. Elevation was used as a representation of soil moisture content; high elevation plots were considered to have low soil moisture levels and low elevation plots to have high soil moisture levels. The purpose of this study was to determine if any synergistic interactions exist between water stress and parasitism on goldenrod floral biomass production. If synergistic interactions did exist between water stress and parasitism, we predicted the summed effects of the simultaneous stressors would be greater than when stressors were present independently. Therefore, we predicted that floral biomass production should be lowest in ramets undergoing gall fly parasitism in high elevation plots (low soil water moisture). Additionally, we wanted to determine if any antagonistic interactions existed between water stress and parasitism on goldenrod ramet height. If antagonistic interactions did exist, the combined effects of both stressors would be less than when each stressor is present independently. Therefore, we predicted that parasitized ramets in high elevation plots (low soil water moisture) would be representative of low water conditions only.
Data was collected on average floral incidences and heights within each elevation and analyzed using ANOVA and interaction plots. Our study was not able to provide data to support that any significant synergistic or antagonistic interactions existed between water stress and parasitism when acting on goldenrod performance. We suspect that the differences in soil moisture between the different elevation sites was not large enough to cause differences in water stress. The inability of our study to support the presence of these relationships does not mean they do not exist in this system and many other terrestrial systems, providing areas for future research. Additionally, the use of natural resources gradients, such as slope, provides an idealistic opportunity to determine the effects of climate change in a natural setting and should be considered for future studies.
Salt: The Element of the Problem
By Sylvia Byer

Sylvia was part of a student group at John F. Ross high school in Guelph, that included Liam Deaton, David Dennis, Charlotte Duncan, Connor Hawkins, Liam Hawkins, Elijah James, Riley Luke, Lucy Matulis, and Ashtyn Yu. The group was supported by science teacher James Bell.
Salt, specifically sodium chloride (NaCl), is used to prevent the formation of ice on roads across Canada. We have observed that the use of NaCl rock salt in Guelph may be excessive, potentially having negative environmental impacts. Our group decided to examine the impact of road salt on white pine trees. We hope that our research can be used to further specific understanding of how NaCl road salt impacts the environment and to highlight the need for less-damaging options for preventing road ice buildup.
A sampling of white pine needles and soil was performed at the Arboretum at the University of Guelph. We collected white pine needles of two different age categories (new needles, old needles) from six trees at three different distances from a salted road (Victoria Road). Soil samples from beneath each of the trees were also collected.
Soil samples were analyzed for soil salinity and pH in the lab of Dr. Longstaffe at the University of Guelph. We measured soil pH using a standard protocol: the soil was suspended in water, allowed to stand, and then pH was measured using a calibrated electrode. Salinity was measured using a similar procedure, except the soil was suspended for 3 minutes to prevent insoluble salts from mixing through reactions.
|
Sample |
Average Salinity (µS/cm) |
Average pH |
|
Closest to the road |
78.43 |
7.54 |
|
Middle |
57.23 |
7.61 |
|
Furthest from the road |
52.40 |
7.65 |
The pine needle samples (new and old) as well as the soil samples taken from beneath the trees were also examined using X-Ray Fluorescence (XRF) at the Canadian Light Source in Saskatchewan. Calcium peaks in both the soil and needles varied with distance from the road, and age of the needles. There was more calcium in the soil closest to the road, however the needles at this distance had the lowest calcium content. Older needles also had a higher calcium content than newer ones.
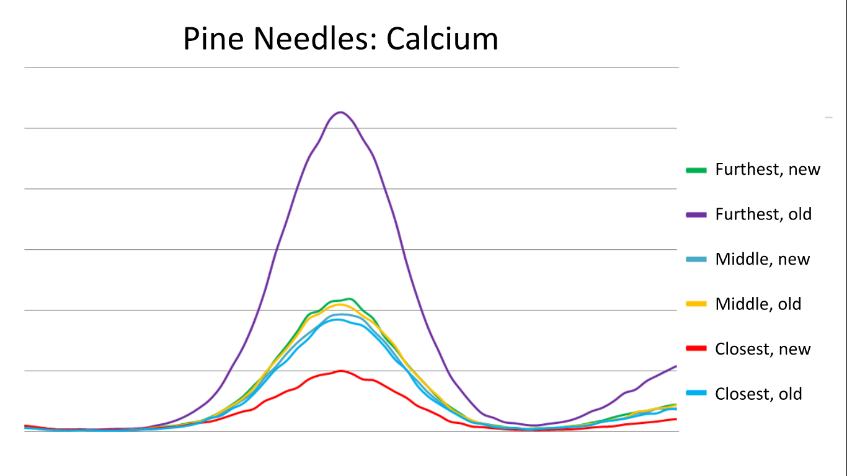
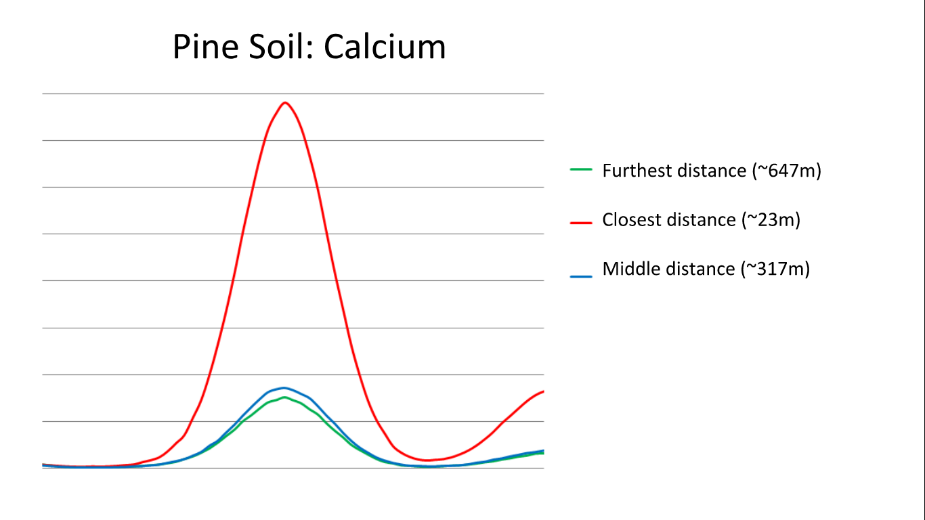
Although our data indicate there was more calcium in the soil close to the road, white pine needles from trees at that location had the least amount of calcium present. It appears the white pine trees were unable to uptake as much of the calcium as trees further from the road despite the higher calcium availability. The results suggest that exposing the trees to salt reduces their ability to uptake calcium, and as calcium is vital to the health of the pine trees, this has the potential to negatively impact the tree. Further research would be needed to determine how severely the lack of calcium is impact the white pine trees.
Building the buzz: Evaluating the efficacy of artificial nest boxes as a conservation tool for bumble bees
By Hayley Tompkins
Hayley Tompkins, a graduate student in Dr. Nigel Raine’s Lab, has been busy at the Arboretum this season installing and monitoring bumble bee nest boxes. Her research, in collaboration with Wildlife Preservation Canada, focuses on the use of artificial nest boxes to study the nesting habits and colony development of wild bumble bees.
Bumble bees are essential pollinators of agricultural crops, and of trees and flowering plants in natural ecosystems. In fact, one in three bites of food we eat depend on the pollination services of insects, such as bumble bees. In Ontario, roughly one third of bumble bee species are in decline, and face threats such as pesticide use, parasites and diseases, and the loss of nesting and overwintering sites because of changes in land use and development.
Not a lot is known about the nesting preferences of bumble bee queens in the spring, such as where different species are nesting and why. To learn more about bumble bee nesting ecology, 82 nest boxes were installed at the Arboretum in mid-April to coincide with the emergence of nest-searching queens. Boxes were installed using one of two techniques (underground and aboveground), and were monitored throughout the spring and summer to look for signs of occupation. During site visits, habitat attributes were collected, and colony development in occupied boxes was monitored. On alternate weeks, site-level floral and bumble bee population surveys were carried out, to collect information on species abundance and distribution.



Nest box setups (left to right): Aboveground domicile, Underground domicile, Occupied Aboveground domicile
Through this research project, Hayley hopes to identify variables leading to successful occupation of nest boxes, and to learn more about colony development of different bumble bee species in Ontario. Preliminary results of this project indicate a 12% occupation rate, which included colonies of at least three different species. The data collected from occupied nest boxes will be used to help fill knowledge gaps in terms of nesting preferences of bumble bee species, and will improve nest box installation techniques in Ontario.
You can view Hayley’s award-winning NSERC “Science, action!” video about her research here: https://www.youtube.com/watch?v=djFDj2hZAj4
Factors affecting senescence in Blue Ash
By Bailey Bingham
Project collaborators: Zach Schonberger, Amy Virostek and Kate Willcox
Fall colours in the Arboretum are not only beautiful but are also very important to the health and success of the trees. The changing colours of the leaves means that the tree’s growing season is over and that they are remobilizing the nutrients in the leaves to other parts of the tree for winter. This process is called autumnal senescence. The timing of autumnal senescence is very important, as a successful tree will keep green leaves for as long as possible (extending its growing season) but remobilize those nutrients and drop its leaves before they are killed by the first frost.
There are two primary mechanisms for how trees determine the best time to change colour and drop their leaves: either it is an evolutionary adaptation (i.e. an internal clock) or it is a plastic response based on local conditions. If it is an adaptation, then trees would be adapted to the photoperiod of their local environment. So, a tree hailing from northern Ontario would be adapted to senescing earlier than a tree from southern Ontario; even if you moved the northern tree to the south, it would still senesce earlier because of its internal clock telling it that winter is coming. If senescence is a plastic response, then a northern tree, moved to the south, might senesce at the same time as its new southern neighbours, because it is responding to the same temperature cues that they are. Because nothing is ever black and white in nature, it is likely that a combination of these two hypotheses is what actually controls senescence. Some researchers have shown that the use of adapted or plastic cues depends on species, while others propose that some trees use both cues, but for different aspects of the senescence process.
For my BIOL*3010 project, I looked at how Blue Ash trees in the Arboretum timed the beginning, the ending and the duration of senescence. Luckily for us, the Arboretum has gene banks which are intended to provide refuge for rare and endangered trees, including Blue Ash (Fraxinus quadrangulata). These gene banks consist of trees grown from seeds obtained from all over Ontario. These collections of trees from different areas, grown in a single location provided a fantastic opportunity to complete our study of the genetic and plastic controls of autumnal senescence.

Our study tracked the senescence process in thirty-four Blue Ash trees from the beginning of September 2017 until the first major frost on October 26, 2017. Overall, our findings supported past literature indicating that different factors may control the different aspects of the process of senescence in these trees. We found that date of onset of senescence was not different between trees from different regions of Ontario, indicating that it may have been a plastic response to local conditions. In comparison, date of completion of senescence was found to vary across trees originating in different townships, which provided support for the idea of an internal clock in these trees.
A limitation of our study was that we couldn’t really look at plastic responses very well, as we only had data from one year. Multi-year analyses would provide a much clearer picture of how individual tree’s timing of senescence responded to yearly variation in temperature.
If any students are interested in continuing this study for your Lab and Field Project, please reach out! It would be really neat to continue this study across multiple years!
Effects of native and non-native predator calls on an alarm behaviour in black-capped chickadees
By Ruby Allen-Powlesland
Project collaborators: Taylor Dupont, Alyssa Robson, Richard Murray, and Nicole Karbusicky
There is increasing interest in integrating discoveries from birds’ anti-predator communication strategies (i.e. alarm calls) into conversation methods. For instance, how might black-capped chickadees (Poecile atricapillus) change the frequency of their infamous ‘chick-a-dee” alarm calls when invasive predators are close? Does the frequency of these alarm calls differ in response to native versus non-native predators?
In our BIOL*3010 research project, we tested the effects of native versus non-native predators’ calls on chickadee alarm behavior. From previous studies, we know that chickadees are able to identify specific predators and warn other flock members in order to defend themselves. Based on this knowledge, we hypothesized that chickadees will have more effective alarm calls in response to established, native predators than in response to newly introduced, non-native predators. This is because chickadees have not learned the predator-specific vocalizations required to identify newer predators to their flock. We predicted that chickadees would respond with more frequent ‘chick-a-dee’ alarm calls in response to native calls than to non-native calls.
To test our hypothesis and predictions, we performed a field study in the University of Guelph Arboretum to observe the calls of chickadees in response to predator auditory cues. Using a speaker to play audio cues, we performed six treatments: three audio recordings of native hawks, which are current predators to chickadees, and three audio recordings of historically absent hawks. We then recorded the number of ‘chick-a-dee’ alarm calls in response.
In our study, chickadees exhibited significantly less ‘chick-a-dee’ calls in response to non-native predator calls than to native predator calls. These results support our hypothesis that chickadees have more effective alarm calls in response to established, native predators than to newly introduced, non-native predators. Further management of non-native predators would benefit chickadee survivorship, since less frequent alarm calling could decrease a chickadee’s ability to warn its flock and protect itself.
Raccoon Latrine Monitoring Project
By Dr. Shannon French, PhD candidate; Department of Pathobiology
Dr. Shannon French, a PhD candidate in the department of Pathobiology is currently investigating the ecology of the raccoon roundworm, Baylisascaris procyonis, an intestinal parasite found in raccoons. One area of her research is investigating the role of rodents and other small animals in maintaining this parasite in the environment.A large number of birds and mammals, including humans, can be infected by the larval stage of the parasite, which is capable of causing neurological disease. These animals, typically referred to as paratenic hosts, become infected by ingesting infective eggs found in raccoon feces. These eggs are resistant to environmental degradation and can persist in the environment for many years. Raccoons become infected either through ingestion of infective eggs, or when they eat an infected bird or rodent.
Raccoons typically use a communal site to defecate, known as a latrine, which serve as the primary source of infection for other animals. Commonly used latrine sites include sheds, attics, fallen logs and other flat elevated surfaces, the base of trees, rocks and tree stumps. Due to the resilience of the parasite eggs, these sites can remain infective even if abandoned and no longer visited by raccoons.



To investigate the transmission of B. procyonis between raccoons and other animals, we are utilizing motion sensing camera traps. In collaboration with the University of Guelph Arboretum, these camera traps are being strategically placed at known raccoon latrines to identify which animals are visiting these sites. This information will help us to determine which animals are at highest risk of exposure to the raccoon roundworm. Preliminary trials have already captured numerous species visiting these sites, including the eastern cottontail, Virginia opossum, eastern chipmunk, and eastern gray squirrel.
A better understanding of the relationship between the raccoon roundworm and its hosts will allow us to better educate the public and make decisions to help prevent exposure to this parasite in the future.
http://blog.healthywildlife.ca/a-study-of-the-raccoon-roundworm-in-ontario-its-a-trap/